|
|
||||||
| >Top >Feature Story >this page | last update:
10/06/30
|
|
| Understanding the immune response mechanism with protein crystallography Scientists at KEK Photon Factory are exploring protein lifecycles—the production, modification, and transportation of proteins—using synchrotron radiation X-ray crystallography. Here, read a story about a regulatory protein called ubiquitin binding to a helical protein NEMO, and how a better understanding of the protein will help drug designers to control immune response in our body. |
|
||||||||||||||||
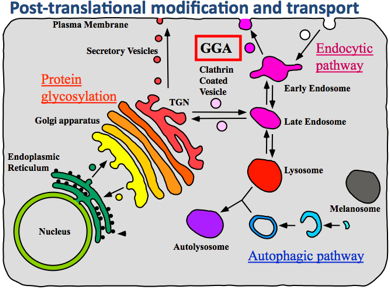
A hundred trillion cells. That is the number of eukaryotic cells in the human body. Each living cell is an enormously elaborate system designed for survival and growth. Each cell houses genetic material, the mechanisms to generate, modify, and transport proteins. Additionally, each cell orchestrates its own metabolism, including cell division, and the lifecycle of proteins. One interesting function of a cell—one which is particularly important to understand if you are a drug designer—is the regulation of responses to external stimuli. The life of a protein molecule starts with protein synthesis. This is the first stage of the protein lifecycle. Here, genetic information in deoxyribonucleic acid (DNA) is copied to produce a ribonucleic acid (RNA) strand. This process is called transcription. The RNA strand then undergoes post-transcriptional modification, where it becomes messenger RNA (mRNA). The mRNA binds to a large protein complex and RNA, called a ribosome, which decodes the mRNA information and creates a protein. This process is called translation. The synthesized protein generally undergoes further modification with other types of proteins, called modification proteins. This is the process of post-translational modification. When the modification is done, proteins are transported to their proper destinations.
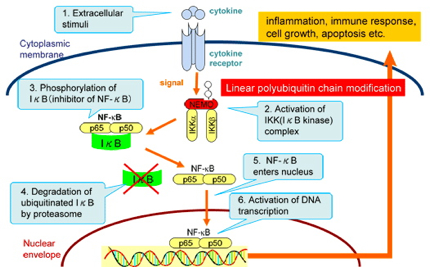
Cells decide the type of proteins to produce depending on their current requirements. In particular, when the cell regulates the immune response to external signals such as viruses, a protein complex called the nuclear factor kappa B, or NF-kappaB, controls the transcription of DNA. NF-kappaB is the key player in regulating the immune response to infectious diseases. The incorrect regulation thereof is linked to tumors, inflammatory diseases, and improper immune developments. and the ubiquitin world
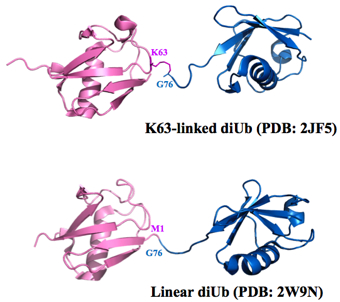
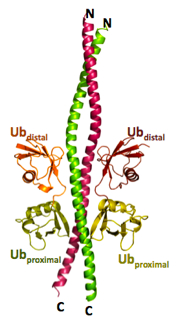
Until recently, only seven types of ubiquitin-ubiquitin bonding were known. The last amino acid in the ubiquitin molecule, the tail, was known to bind to any of the seven lysines—6th, 11th, 27th, 29th, 33rd, 48th, or 63rd amino acids—in another ubiquitin molecule. Then, in 2008, a group from Osaka University discovered a new type of connection which they called the linear chain. In a linear chain, the head of one ubiquitin molecule bonds with the tail of another. The number of links in a ubiquitin chain could be, theoretically, any number. With the addition of the head-to-tail linear linkage, the number of possible combinations of multiple ubiquitins linked quickly grew. Approximately 27,000 papers have already been published on ubiquitin so far. “The ubiquitin world is new, and the horizon is wide open and far away,” says Wakatsuki. Finding NEMO… with ubiquitin While exploring the ubiquitin world, Dr. Simin Rahighi of KEK, a postdoc at Wakatsuki’s laboratory, made several major findings, eventually uncovering the biological importance of the linear di-ubiquitin molecule. Through a combination of hard work, and a little bit of chance, Rahighi was able to develop a very clear picture of the NF-kappaB essential modifier (NEMO) binding to a linear di-ubiquitin, in which two ubiquitin molecules are connected by a head-to-tail bond. This structure of the NEMO-di-ubiquitin complex amazed the community and also stirred up a controversy.
The strand of ubiquitin is folded up into a very small volume in a particular way. It happens that, in this folded configuration, the head of the protein is very close to the K63 amino acid (‘K’ stands for lysine, while the number stands for the address of the amino acid). Because of the geometrical proximity, the shape of the head-to-tail linear di-ubiquitin is very similar to that of the K63 linked di-ubiquitin. Since protein function is strongly influenced by protein shape, many people, including Wakatsuki, thought that the biological function of these two di-ubiquitin structures would be very similar.
On the other hand, a di-ubiquitin with linear linkage was much simpler to synthesize. The head-to-tail structure could be created through simple genetic engineering. Instead of linking after the protein synthesis, the two DNA sequences of ubiquitin are first connected before the protein synthesis so that it produces a linearly linked di-ubiquitin. “Many years of development in genetic engineering now allow us to freely change DNA sequences,” says Wakatsuki. “In fact, most of proteins used in biological experiments are now produced by genetic engineering.”
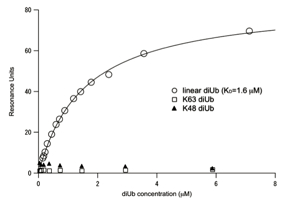
With this new strategy, Rahighi set off to see how a linear di-ubiquitin would interact with NEMO. The first step was a test using a measurement method called surface Plasmon resonance (SPR), which measures the degree of resonance between two molecules. The results surprised everyone. The resonance curves showed that linear di-ubiquitin strongly binds to NEMO, while the K63 linked di-ubiquitin did not. Crystallizing the ubiquitin-NEMO complex Excited by these results, Rahighi began trying to grow crystals of NEMO in order to discover the structure of NEMO. This was in October 2006. Just six months later, she was able to solve the NEMO structure. Next, she launched into a new project: growing crystals of the NEMO and linear di-diubiquitin complex. Growing clean, flawless crystals with exact periodic structure is the key to crystallography, and yet it is very difficult with small, dynamic biomolecules. Rahighi tested a hundred different detergent additives, and hundreds of different growing conditions. Eventually, she succeeded, and brought her crystals to the KEK Photon Factory. There, she shot the crystals with X-ray beam, and examined the resulting diffraction patterns. “I was able to get good diffraction data that were supposed to be sufficient for my analyses,” says Rahighi.
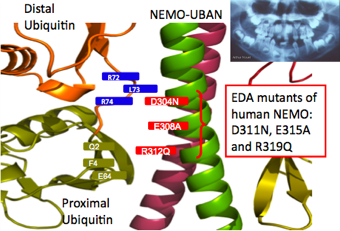
“The approach was basically try-and-error,” says Rahighi. “It was very frustrating. I knew I was so close to the goal, yet it was still so far.” In her experiment, she tried introducing heavy atoms into protein crystals, got diffractions, but signals from heavy atoms were not sufficient for phasing. She also tried mutating her protein complex in various ways, none of which worked for her until she came across one rather uncommon approach: surface entropy reduction (SER). SER is an experimental method, in which some of the amino acid residues located at the surface of the protein are mutated to lower the surface entropy. In particular, the residues in NEMO proteins that have long side chains are mutated to shorter ones. This in turn reduces the entropy—randomness—of the system, making it easier to crystallize. With SER, the structure of the resulting NEMO-ubiquitin complex could be solved, and showed a clear binding between two linearly linked di-ubiquitins and the NEMO molecule. Up to this point, scientists knew that NEMO bind to linear ubiquitin chains much stronger than to K63 linked ubiquitin chains, but did not know why. The crystallography gave a definitive answer to this question. The most accurate NEMO-ubiquitin structure At first, Rahighi’s finding was met with skepticism. A model of NEMO-ubiquitin structure had already been published by a group at the Weill Cornell Medical College in the journal Molecular Cell in January 2009. Based on nuclear magnetic resonance (NMR) spectroscopy, they believed the NEMO-ubiquitin structure consisted of a single di-ubiquitin binding to one NEMO dimer.
Rahighi's results can be applied to drug design. For example, when an immune response in our body is causing damage, as is the case with hay fever, one can suppress the level of response by cutting away the linear ubiquitin chain from NEMO in only the area of inflammation. “In drug design, drug delivery to the target area is extremely important,” says Wakatsuki. “We also study genetic modification of protein transport to bring about speedy drug delivery.”
At the KEK Photon Factory, a new beamline BL-1A just began operation last month, producing brilliant, low-energy X-ray beam. This enables structural biology scientists to solve the phase problem using anomalous signals from sulfur atoms in natural protein crystals. Rahighi continues to explore the ubiquitin world with the new beamline. |
| copyright (c) 2010, HIGH ENERGY ACCELERATOR RESEARCH ORGANIZATION, KEK 1-1 Oho, Tsukuba, Ibaraki 305-0801 Japan |
||